An advanced course on bacterial translation and physiology is to be taught at the RiboCORE, Uppsala University in spring 2013. I will be giving a lecture on the stringent response. You can find the course programme here. Review on the stringent response written by Gem Atkinson and myself is a nice introduction to the subject.
Wednesday, December 19, 2012
Thursday, November 15, 2012
GDP and SRL don't mix
Translational GTPases run the ribosomal cycle, and the ribosome talks back - it recruits the GTPases when it is a certain state, affects trGTPase's affinity to G nucleotides and activates the GTP hydrolysis when needed. Using Isothermal Titration Calorimetry we showed that binding of GDP nucleotide and of SRL rRNA element to translational GTPases IF2 and EF-G are mutually exclusive. This suggests a neat mechanism for the destabilisation of the ribosome-bound GDP form of the GTPase: the moor has done his duty, the moor can go.
Due to the technical limitations, the ITC experiments were performed with a 27-nucleotide long RNA piece mimicking the rRNA element as a model. In order to place our results in the framework of the ribosomal cycle we need experiments with the whole ribosome.
References:
Mitkevich et al., Scientific Reports 2012 2:843, PIMD: 32150791
Due to the technical limitations, the ITC experiments were performed with a 27-nucleotide long RNA piece mimicking the rRNA element as a model. In order to place our results in the framework of the ribosomal cycle we need experiments with the whole ribosome.
References:
Mitkevich et al., Scientific Reports 2012 2:843, PIMD: 32150791
Wednesday, November 7, 2012
First PhD defence in the lab
Our first PhD defence took place on November 2d, 2012. Viktoriya Shyp has defended her work "G nucleotide regulation of translational GTPases and the stringent response factor RelA". Mike Cashel, the discoverer of ppGpp, served as opponent.
Hurray to Vika!
Hurray to Vika!
Wednesday, October 3, 2012
Relacin - a novel antibacterial targeting the stringent response, maybe
The stringent response is a promising target for novel antibacterials: it is involved in virulence and antibiotic insensitivity, and inhibiting the stringent response would disarm the bug, making is both less evil and easier to kill.
A new study is came out in PLoS Pathogens describing a novel Rel inhibitor, relacin (Fig. 1). Wexselblatt and colleagues are following up their earlier work on derivatizing ppGpp into a Rel inhibitor and are now testing the compound not only in vitro, but also in vivo.

Fig 1: the chemical structure of relacin.
They show that relacin efficiently inhibits sporulation of Bacillus subtilis. Sporulation in this organism is driven by ppGpp, and inhibitory effect of relacin is a strong indication that it actually works. However, really high concentrations are needed to achieve significant effects: 0.5 - 2 mM. At these concentrations one would expect that in addition to hitting RelA, relacin will affect all the other ppGpp targets, i.e. translational GTPases, GTP biosynthesis enzymes etc. The authors do not test these effects. It would be easy to do it in an in vitro translational lysate... but, unfortunately, this is not done. By using a GFP-fusion reporter, they do show that relacin inhibits translation of mid-sporulation protein SpoIIQ, but they do not check that it does not inhibit translation in general. A simple test of GFP expression would do.
With this (potential) absence of specificity relacin is unlikely to be the 'magic bullet' inhibiting just the stringent response and making bacterial less pathogenic, but still viable. However, relacin is just the first step. There is a hope that the derivatives to come will work at more in vivo-relevant concentrations and will be highly Rel-specific.
References:
Wexselblatt et al. Biomed Org Chem (2010) PIMD: 20483622
Friday, September 14, 2012
ppGpp regulates GTP synthesis by inhibiting Gmk and HprT
Another role for ppGpp was discovered by Allison Kriel and colleagues: it directly regulates GTP levels by interfering with GTP biosynthetic pathway. It is worth saying that inhibitory effects of (p)ppGpp on IMP dehydrogenase and anenylosuccinate synthetase, first enzymes of the guanylate and adenylate pathways, respectively, were discovered by Gallant and colleagues as early as in 1971, so the connection between (p)ppGpp and metabolism of G nucleotides was known long ago.
The cool thing about the paper is the approach they use. There can be loads of potential targets of ppGpp, and loads of proteins will be inhibited by it in vitro. Which ones are the relevant ones? Kriel and colleagues usa a top-down approach: first they get a birds-eye view of starvation by running metabolic and transcriptomic analysis of starved wt and ppGpp0 (i.e. devoid of ppGpp) strains, then identify the ppGpp targets by clustering and pathway analyses, and then follow their predictions up in vitro and an vivo. This is a really powerful approach.
The net result is that (p)ppGpp inhibits several enzymes in the GTP biosynthesis pathway, and by doing quantitative experiments, Kriel et al. identify the primary targets, Gmk and HprT (Fig. 1). All the experiments are done in B. subtilis, and in this bug pppGpp is the major magic spot nucleotide. It is made of GTP, so inhibition of GTP synthesis results in a negative feedback control loop. This loop turns out to be a key component for the control of the GTP levels in the cell.
In B. subtilis, unlike E. coli, ppGpp does not regulate RNA polymerase directly: the regulation goes via effects on the GTP level. And indeed, a genetic screen performed by Kriel et al. showed that regulation of GTP metabolises by ppGpp, not of polymerase is crucial for the bacterial survival under stress. The have found 37 suppressor mutations leading to survival of the ppGpp0 strain - and most of these turned out to be in the GTP biosynthesis pathway. In the E. coli case, the supressor mutations are usually in the RNA polymerase.
The negative control of GTP synthesis by ppGpp turned out to be crucial for bacterial well-being - in the ppGpp0 cells high levels of GTP caused cell death, though the mechanism is still unclear (Fig. 1). Kriel et al. proposed several possible explanations: inhibition with ATP-consuming enzymes, excessive up-regulation of the rRNA transcription, effects on dGTP synthesis etc.

Fig. 1: ppGpp's role in B. subtilis survival via regulation of GTP biosynthesis. Figure from Kriel et al.
References:
Kriel et al, Cell (2012) in press
Gallant et al. JBC (1971), 246 (18): 5812-5816, PIMD: 4938039
Krasny and Grouse, EMBO J. (2004), 23(22): 4473-83, PIMD: 15496987
The cool thing about the paper is the approach they use. There can be loads of potential targets of ppGpp, and loads of proteins will be inhibited by it in vitro. Which ones are the relevant ones? Kriel and colleagues usa a top-down approach: first they get a birds-eye view of starvation by running metabolic and transcriptomic analysis of starved wt and ppGpp0 (i.e. devoid of ppGpp) strains, then identify the ppGpp targets by clustering and pathway analyses, and then follow their predictions up in vitro and an vivo. This is a really powerful approach.
The net result is that (p)ppGpp inhibits several enzymes in the GTP biosynthesis pathway, and by doing quantitative experiments, Kriel et al. identify the primary targets, Gmk and HprT (Fig. 1). All the experiments are done in B. subtilis, and in this bug pppGpp is the major magic spot nucleotide. It is made of GTP, so inhibition of GTP synthesis results in a negative feedback control loop. This loop turns out to be a key component for the control of the GTP levels in the cell.
In B. subtilis, unlike E. coli, ppGpp does not regulate RNA polymerase directly: the regulation goes via effects on the GTP level. And indeed, a genetic screen performed by Kriel et al. showed that regulation of GTP metabolises by ppGpp, not of polymerase is crucial for the bacterial survival under stress. The have found 37 suppressor mutations leading to survival of the ppGpp0 strain - and most of these turned out to be in the GTP biosynthesis pathway. In the E. coli case, the supressor mutations are usually in the RNA polymerase.
The negative control of GTP synthesis by ppGpp turned out to be crucial for bacterial well-being - in the ppGpp0 cells high levels of GTP caused cell death, though the mechanism is still unclear (Fig. 1). Kriel et al. proposed several possible explanations: inhibition with ATP-consuming enzymes, excessive up-regulation of the rRNA transcription, effects on dGTP synthesis etc.

Fig. 1: ppGpp's role in B. subtilis survival via regulation of GTP biosynthesis. Figure from Kriel et al.
References:
Kriel et al, Cell (2012) in press
Gallant et al. JBC (1971), 246 (18): 5812-5816, PIMD: 4938039
Krasny and Grouse, EMBO J. (2004), 23(22): 4473-83, PIMD: 15496987
Thursday, September 13, 2012
One more GTPase that binds ppGpp: RF3
The stringent response alarmone ppGpp is very, very similar to GDP: add two more phosphates and you have it... So it is only natural that GTPases mistake the two.
Now Kihira and collegues has added one more to the list - bacterial termination factor 3, RF3. This factor is involved in release of the class one release factors, RF1 and RF2. It binds to the ribosome in the GDP-bound state, exchanges GDP to GTP and kicks the class 1 factors off. ppGpp:RF3 complex is not as active as GDP:RF3 one, a situation similar to that with initiation factor 2, IF2.
The question is is it a specific regulatory mechanism or are GTPases just promiscuous enough? After all, even archael elongation factor 1A, aEF1A, is inhibited by ppGpp as well. And these two never meet in nature...
Kanjee and collegues have recently compiled a list of proteins that ppGpp binds to... or is expected to bind to. GTPases are there, as a whole class of enzymes.
References:
Kihira et al. FEBS J. (2012) in press
Kanjee et al. Mol. Microbiology (2012): 85 (6) 1029-1043 PIMD: 22812515
Now Kihira and collegues has added one more to the list - bacterial termination factor 3, RF3. This factor is involved in release of the class one release factors, RF1 and RF2. It binds to the ribosome in the GDP-bound state, exchanges GDP to GTP and kicks the class 1 factors off. ppGpp:RF3 complex is not as active as GDP:RF3 one, a situation similar to that with initiation factor 2, IF2.
The question is is it a specific regulatory mechanism or are GTPases just promiscuous enough? After all, even archael elongation factor 1A, aEF1A, is inhibited by ppGpp as well. And these two never meet in nature...
Kanjee and collegues have recently compiled a list of proteins that ppGpp binds to... or is expected to bind to. GTPases are there, as a whole class of enzymes.
References:
Kihira et al. FEBS J. (2012) in press
Kanjee et al. Mol. Microbiology (2012): 85 (6) 1029-1043 PIMD: 22812515
Monday, July 30, 2012
Positive feedback control of E. coli RelA by its product ppGpp
ppGpp regulates numerous targets, and now we added one more: the stringent response factor RelA itself. Using an in vitro stringent response system we showed that ppGpp dramatically increases the turnover rate of RelA, both is the system where RelA is activated by the ribosomes (both naked and programmed with tRNA and mRNA) and in the system where RelA is activated by the ribosomal protein L11 alone.
 Figure 1: RelA activation in the 70S-driven in vitro system upon addition of ppGpp
Figure 1: RelA activation in the 70S-driven in vitro system upon addition of ppGpp
We did 70S and L11 tittrations and demonstrated that ppGpp increases RelA's kcat, making it a more efficient enzyme:

Figure 2. RelA activity as a function of the 70S concentration in presence and absence of ppGpp
What next? First off, we do not know where ppGpp binds and how it regulates RelA on the mechanistic level. Second, since there are at least 30 groups of the RSH proteins, we will figure out which are activated by this mechanism, and which are not. This will provide us some vital clues for understanding the computational properties of the stringent response system. Third, after this in vitro result it is instrumental to show the ppGpp-mediated activation in vivo.
PS: and now our paper got covered as a Research Highlight in Nature Chemical Biology. Yay!
References:
Shyp et al., EMBO Reports (2012) doi: 10.1038/embor.2012.106.
PIMD: 22814757

We did 70S and L11 tittrations and demonstrated that ppGpp increases RelA's kcat, making it a more efficient enzyme:
Figure 2. RelA activity as a function of the 70S concentration in presence and absence of ppGpp
What next? First off, we do not know where ppGpp binds and how it regulates RelA on the mechanistic level. Second, since there are at least 30 groups of the RSH proteins, we will figure out which are activated by this mechanism, and which are not. This will provide us some vital clues for understanding the computational properties of the stringent response system. Third, after this in vitro result it is instrumental to show the ppGpp-mediated activation in vivo.
PS: and now our paper got covered as a Research Highlight in Nature Chemical Biology. Yay!
References:
Shyp et al., EMBO Reports (2012) doi: 10.1038/embor.2012.106.
PIMD: 22814757
Monday, July 9, 2012
iGEM2011: GFP-based readout for ppGpp concentration in vivo
Measuring ppGpp concentration in individual living cells with good temporal resolution would be great. I've been musing about a possibility of doing that using RNA aptamers, but that's just musing. It seems like am iGEM team from the University of Trondheim tried setting up a GFP-based reported system, and this system is, maybe, possibly, probably, working. Somewhat.
Unfortunately there is no publication. However, there is a popular article about the whole affair, in Norwegian and there is a short report on the iGEM webpage.
Apropos to the technical issues that are listed in the original report, such as dramatic leakage of the GFP reporter in the absence of stress stimuli, there are several conceptual concerns. First, the system is based on translation of the GFP reporter during the stringent response, and during the stringent response translation is strongly inhibited. Second, GFP (they use red version of it, mCherry) has to mature in order to become bringt, and that takes some time - from minutes to hours, depending on the conditions and what sort of GFP variant it is. For mCherry maturation time is 15-40 minutes, and this is comparable with E. coli generation time. Therefore, first, one would expect a very pronounced lag before the SR is engaged and the corresponding readout and, second, all the fluctuations in the ppGpp concentrations happening on the timescale below 10s of minutes will be averaged out. Third, GFP is very stable, so this reporter system will have severe memory effects - once the cell has committed to stringency, it will produce GFP, and even though stringency is reversed, GFP will stay. Maybe it is possible to turn this into a feature, but I am not sure how. And, lastly, brightness of the GFP depends on the pH and redox potential of the environment, and these things change in the stressed cells.
Unfortunately there is no publication. However, there is a popular article about the whole affair, in Norwegian and there is a short report on the iGEM webpage.
Apropos to the technical issues that are listed in the original report, such as dramatic leakage of the GFP reporter in the absence of stress stimuli, there are several conceptual concerns. First, the system is based on translation of the GFP reporter during the stringent response, and during the stringent response translation is strongly inhibited. Second, GFP (they use red version of it, mCherry) has to mature in order to become bringt, and that takes some time - from minutes to hours, depending on the conditions and what sort of GFP variant it is. For mCherry maturation time is 15-40 minutes, and this is comparable with E. coli generation time. Therefore, first, one would expect a very pronounced lag before the SR is engaged and the corresponding readout and, second, all the fluctuations in the ppGpp concentrations happening on the timescale below 10s of minutes will be averaged out. Third, GFP is very stable, so this reporter system will have severe memory effects - once the cell has committed to stringency, it will produce GFP, and even though stringency is reversed, GFP will stay. Maybe it is possible to turn this into a feature, but I am not sure how. And, lastly, brightness of the GFP depends on the pH and redox potential of the environment, and these things change in the stressed cells.
Thursday, July 5, 2012
More fun with fun12... right, that was a dreadful joke
I like translational GTPases. I particularly like bacterial ones. Hell, I am titrating one with GTP as we speak, so I am partial. However, eukaryotic translational GTPases are also OK, particularly the ones that have bacterial homologues.
One of these is eukaryotic initiation factor 5B, eIF5B, a homologue of bacterial initiation factor 2, IF2. In S. cerevisiae eIF5B is called Fun12. eIF5B is a GTPase, and translational GTPases are tightly regulated. During translation initiation Fun12 is involved in the subunit joining, and GTP hydrolysis is coupled with factor's release from the initiation complex.
Now it turnes out that Fin12/eIF5B has another function outside of translation initiation: together with the 60S subunit it proofreads the ribosomal assembly (Lebaron et al. 2012 and Strunk et al. 2012). Assembly of the ribosomal subunits is a multistep process, with r-proteins binding is certain sequence, assisted by various assembly factors, with rRNA being cut and remodeled. One of the steps is cleavage of the 18S rRNA by the Nob1 RNAse. And this stage, apparently, is stimulated by Fun12. Bacterial Fun12 homologue, IF2, is GTP-dependently rearranging the ribosomal structure by inducing intersubunit rotation, so the same process is now suggested to play the role in 40S maturation in yeast via driving the ribosome in Nob1-susceptible state.
In eucaryotes translation-incompetent ribosomes undergo so-called nonfunctional rRNA decay, NRD. By linking functionality on Fun12 binding and activation via consecutive 60S binding, translational functionality is linked to rRNA processing, providing another safety net making sure that only functional ribosomes are involved in translation, and the nonfunctional ones are rapidly cleared out.
In eucaryotes translation-incompetent ribosomes undergo so-called nonfunctional rRNA decay, NRD. By linking functionality on Fun12 binding and activation via consecutive 60S binding, translational functionality is linked to rRNA processing, providing another safety net making sure that only functional ribosomes are involved in translation, and the nonfunctional ones are rapidly cleared out.
References:
Lebaron et al. Proofreading of pre-40S ribosome maturation by a translation initiation factor and 60S subunits. Nature Str. Mol. Biol. 2012 in press PIMD: 22751017
Strunk et al. A Translation-Like Cycle Is a Quality Control Checkpoint for Maturing 40S Ribosome Subunits. Cell 2012 vol. 150 (1) pp. 111-121 PIMD: 22770215
Tuesday, July 3, 2012
One more '-omics' analysis of the stringent response: BIBLIOMICS
The stringent response is confusing, no doubt about that. I personally get exceedingly confused when I read in vivo papers from the 80s. I somehow hope that there should be hidden gems there, and it is just my stupidity that stops me from discovering these.... so I try... and I get confused. In vivo data confuse me, and in vivo data from the 80s.... I am lost.
Importantly, I always try to read one paper at a time, maximum ten, rarely more then twenty. Imagine what happens if one would read them all? And how, I wonder, how would one call this sort of thing? Wonder no more; enter Carneiro and colleagues. What they did, they collected the whole bibliome about the E. coli stringent response and analyzed it in attempt to gain a birds-eye view, providing 'a more systematic understanding of this cellular response'.
They summarize the nitty-gritty of the stringent response in the magnificent Figure 1.

The bird-eye view allows Carneiro and collegues to make some information-packed generalizations. I am not sure what we learn this way, but the figures speak for themselves:

Rrrright, the dashed line overtook the solid one and they never crossed, therefore the ratios between the blue and red bars changed with time... I want some error-bars, then it will be really, really nice and scientific.
But it is not only the figures that are great. The text is awesome as well. 'Later, in 1980, the ppGpp level was found to be controlled by the SpoT enzyme via GTP hydrolysis activity (PMID: 6159345)'. GTP hydrolysis! By SpoT! Yay! 'The 50S ribosomal subunit protein L11 has been indirectly implicated in the feedback inhibition of (p)ppGpp, because ribosomes lacking this protein are unable to stimulate the synthesis of these nucleotides (PMID:11673421; PMID:17095013) [39,61].' Feedback inhibition! This redefines the meaning of feedback... Wow! No wonder L11 migrated on the large subunit...
Being subjected to such a monumental degree of confusion, the authors started expressing themselves in a most peculiar way; for the lack of a better word I would call it 'cautious': 'As a result, it was possible to perceive the relevance of specific translation GTPases known to be inhibited by (p)ppGpp nucleotides'. 'Studies showed that (p)ppGpp inhibits translation by repressing the expression of ribosomal proteins and also potentially inhibiting the activity of the particular proteins'.
I certainly hope that soon the authors should take on the mighty ribosome. There are many more papers in the ribosomal bibliome, and the level of confusion might be even higher. I would also recommend some 3D plots!
References:
Carneiro, S., Lourenço, A., Ferreira, E. C., & Rocha, I. (2011). Stringent response of Escherichia coli: revisiting the bibliome using literature mining. Microbial informatics and experimentation, 1(1), 14. doi:10.1186/2042-5783-1-14 PIMD: 22587779
One more crazy little thing
Stringent response is run by a family of proteins called RelA-SpoT Homologues, RSH, and these come in two flavors: the long ones and the short ones. The long ones have both ppGpp synthesizing and ppGpp hydrolyzing domains, and either both are active, with synthesizing being the dominant one (that would be the ancestral Rel) or both are active, with hydrolyzing being the dominant one (SpoT) or only the synthesizing one is active (RelA). The short RSHs are more variable. They have only one of the domains, so they can either synthesize (SAS, small alarmone synthetase) or hydrolyze (SAH, small alarmone synthetase) ppGpp. What is fun, is that in addition they can have other domains, sometimes with very peculiar functions.
A very peculiar SAS was characterized recently by Maya Murdeshwar and Dipankar Chatterji. They call it MS_RHII-RSD, or, using terminology proposed in our paper with Gemma Atkinson, actRelMsm. This SAS from Mycobacterium smegmatis in addition to the ppGpp synthetic activity has another one, quite unexpected. It has a dedicated domain capable of hydrolyzing DNA:RNA duplexes via its RNAse H domain.
Quite bizarre, quite.
References:
MS_RHII-RSD: a dual function RNase HII - (p)ppGpp synthetase from Mycobacterium smegmatis. M. Murdeshwar and D. Chatterji. J. Bacteriology, 2012, in press PIMD: 22636779
References:
MS_RHII-RSD: a dual function RNase HII - (p)ppGpp synthetase from Mycobacterium smegmatis. M. Murdeshwar and D. Chatterji. J. Bacteriology, 2012, in press PIMD: 22636779
Wednesday, March 14, 2012
Proline residue in L11 as a key regulator of translational GTPases?
The ribosome is run by translational GTPases. Translational GTPases, in their turn, are regulated by the ribosome. They all bind in the same region (GAC, GTPase associated center) of the ribosome. In bacteria the GTPase binding site consists of a couple of rRNA elements: SRL (sarcin-ricin loop) and thiostrepton loop and several ribosomal proteins:L7/L12 stalk (L10 and L7/L12) and L11.
The latter is the main hero of a fresh paper in Nature Structural and Molecular Biology by Wang and coworkers. They show that bacterial translational GTPases (such as EF-G) when binding to the ribosome act as peptidyl-prolyl cis-trans isomerases (PPIases) driving isomerisation in the conserved residue in the ribosomal protein L11. This isomerisation, in turn, transmits signal to the ribosomal protein L7 /L12 - something that is necessary for efficient GTPase cycling on the ribosome.
I like L11 - it is a key protein for stringent response, and without it stringent factor RelA does not work, as was shown using E. coli mutants lacking L11 (Dabbs J. Bac 1979). These mutants are perfectly viable, but grow ten times slower then the wild type E. coli, most probably due to defects in the ribosome assembly (Hampl et al. JBC 1981). The very viability of the L11 knock-out strains tells us that L11 is not the key for keeping the ribosome running. In fact, less than a half of ribosomal proteins can be knocked-out in E. coli (22 out of 54, Shoji et al. JMB 2011), making L11 one of the less-important ones... and keeping an eye of the translational GTPases is definitely not one of the less-important functions!
This seems to be bit paradoxical - a ribosomal protein that is dispensable involved in something that is very central for protein biosynthesis. It gets even more fascinating when you look at the evolutionary aspect of the story (Gem Atkinson does that in her blog). Wang and colleagues managed to map the PPIase site of EF-G. As they show PPIase activity is universal for all the bacterial translational GTPases they tested, and the PPIase site is, surprisingly, quite a variable region of the G domain! So, do they all reinvent the weel separately? This is all most peculiar.
References:
Wang et al. A conserved proline switch on the ribosome facilitates the recruitment and binding of trGTPases. Nat Struct Mol Biol (2012) PIMD: 22407015The latter is the main hero of a fresh paper in Nature Structural and Molecular Biology by Wang and coworkers. They show that bacterial translational GTPases (such as EF-G) when binding to the ribosome act as peptidyl-prolyl cis-trans isomerases (PPIases) driving isomerisation in the conserved residue in the ribosomal protein L11. This isomerisation, in turn, transmits signal to the ribosomal protein L7 /L12 - something that is necessary for efficient GTPase cycling on the ribosome.
I like L11 - it is a key protein for stringent response, and without it stringent factor RelA does not work, as was shown using E. coli mutants lacking L11 (Dabbs J. Bac 1979). These mutants are perfectly viable, but grow ten times slower then the wild type E. coli, most probably due to defects in the ribosome assembly (Hampl et al. JBC 1981). The very viability of the L11 knock-out strains tells us that L11 is not the key for keeping the ribosome running. In fact, less than a half of ribosomal proteins can be knocked-out in E. coli (22 out of 54, Shoji et al. JMB 2011), making L11 one of the less-important ones... and keeping an eye of the translational GTPases is definitely not one of the less-important functions!
This seems to be bit paradoxical - a ribosomal protein that is dispensable involved in something that is very central for protein biosynthesis. It gets even more fascinating when you look at the evolutionary aspect of the story (Gem Atkinson does that in her blog). Wang and colleagues managed to map the PPIase site of EF-G. As they show PPIase activity is universal for all the bacterial translational GTPases they tested, and the PPIase site is, surprisingly, quite a variable region of the G domain! So, do they all reinvent the weel separately? This is all most peculiar.
References:
Tuesday, March 13, 2012
ppGpp induces production of fruiting bodies in Myxococcus xanthus
E. coli is boring, admit it. At least in comparison with Myxococcus xanthus: a self-organized, predatory saprotrophic single-species biofilm called a swarm according to the Wikipedia. Now that sounds exciting! I wish one day somebody would call me "a self-organized, predatory biofilm called a swarm"! That would make a lovely email signature: "Vasili Hauryliuk, PhD, self-organized, predatory biofilm called a swarm". Hell yes.
Harris BZ, Kaiser D, & Singer M (1998). The guanosine nucleotide (p)ppGpp initiates development and A-factor production in Myxococcus xanthus. Genes & Development, 12 (7), 1022-35 PMID: 9531539
But I digress. Stringent response (or, to be more specific, RelA-mediated production of alarmone molecule ppGpp) regulates loads of things in bacterial physiology: it turns on bacterial survival mode and shuts down production of ribosomes, it induces virulence (cornered bacteria are deadly) and makes bugs more resistant to antibiotics. Now let us just imagine for a moment what stringent response can do to a "a self-organized, predatory biofilm called a swarm"! Exactly that was investigated in recent paper by Konovalova and colleagues (Konovalova et al. Mol Microbiology 2012).
Unlike boring E. coli, Myxococcus xanthus has a life cycle (Fig. 1). It can swarm happily gobbling up other bacteria, or, if food supply is low, it can form a fruiting body (a life-stile similar to that of slime molds who are not bacteria but eucaryotes).

Fig. 1. Life cycle of a self-organized, predatory biofilm called a swarm (AKA Myxococcus xanthus).
Formation of the fruiting bodies depends on the functionality of the stringent response system (Harris et al. Gens Dev. 1998). How Konovalova and colleagues fill in the molecular details presenting an example of post-translational activation of secretion by regulated proteolysis. Here is how it works.
Formation of the fruiting bodies depends on the cell-to-cell signaling, and this process, obviously, happens outside of the cell. It involves proteolysis of several extracellular target proteins by a subtilisin-like protease PopC, which needs to be exported outside of the cell in order to do its job. So now it turns out that RelA, working together with PopD protein, regulates PopC export, which is activated during starvation (and, therefore, production of ppGpp). The PopD:PolC complex formation is not affected by ppGpp, suggesting that regulation of export by RelA is using some indirect mechanism. And indeed, PopD turned out to be degraded during starvation in a FtsH-dependent manner, releasing PopC - a story somewhat similar to regulation of toxin:antitoxin pairs via antitoxin degradation by Lon protease during nutritional stress.
All this brings us to the question of importance of the regulated proteolysis during the stringent response. One known example of ppGpp-mediated control via protein degradation is degradation of ribosomal proteins by Lon protease induced by accumulation of polyphosphate. Unfortunately, usually stringent response on the whole-cell level is studied on the mRNA level, by, say, microarrays. It would be most educational to compare the changes on the mRNA level with changes on the proteome level and to pick up the protein degradation-mediated regulation pathways.
References:
Unlike boring E. coli, Myxococcus xanthus has a life cycle (Fig. 1). It can swarm happily gobbling up other bacteria, or, if food supply is low, it can form a fruiting body (a life-stile similar to that of slime molds who are not bacteria but eucaryotes).

Fig. 1. Life cycle of a self-organized, predatory biofilm called a swarm (AKA Myxococcus xanthus).
Formation of the fruiting bodies depends on the functionality of the stringent response system (Harris et al. Gens Dev. 1998). How Konovalova and colleagues fill in the molecular details presenting an example of post-translational activation of secretion by regulated proteolysis. Here is how it works.
Formation of the fruiting bodies depends on the cell-to-cell signaling, and this process, obviously, happens outside of the cell. It involves proteolysis of several extracellular target proteins by a subtilisin-like protease PopC, which needs to be exported outside of the cell in order to do its job. So now it turns out that RelA, working together with PopD protein, regulates PopC export, which is activated during starvation (and, therefore, production of ppGpp). The PopD:PolC complex formation is not affected by ppGpp, suggesting that regulation of export by RelA is using some indirect mechanism. And indeed, PopD turned out to be degraded during starvation in a FtsH-dependent manner, releasing PopC - a story somewhat similar to regulation of toxin:antitoxin pairs via antitoxin degradation by Lon protease during nutritional stress.
All this brings us to the question of importance of the regulated proteolysis during the stringent response. One known example of ppGpp-mediated control via protein degradation is degradation of ribosomal proteins by Lon protease induced by accumulation of polyphosphate. Unfortunately, usually stringent response on the whole-cell level is studied on the mRNA level, by, say, microarrays. It would be most educational to compare the changes on the mRNA level with changes on the proteome level and to pick up the protein degradation-mediated regulation pathways.
References:
Konovalova A, Löbach S, & Søgaard-Andersen L (2012). A RelA-dependent two-tiered regulated proteolysis cascade controls synthesis of a contact-dependent intercellular signal in Myxococcus xanthus. Molecular Microbiology PMID: 22404381
Double life of mitochondrial ribosomal protein L7 12
Mitochondria have their own transcriptional and translational apparatus, even though they produce only a handful of proteins, therefore most of the proteins are imported from the cytoplasm. Trancription, translation and protein insertion into the membrane are interconnected: translational activators regulating mitochondrial translation are interacting with mitochondrial RNA polymerase via Nam1p and Sls1p proteins (Bryan et al. Genetics 2002), Puf proteins connect cytoplasmic translation and protein import into mitochondria by direct interaction with Tom20 subunit of the TOM protein import channel (Saint-Georges et al. PLoS ONE 2008).
But this seems not tight enough interaction for mitochondrial translation and transcription. It turnes out what mitohondrial ribosomal protein L7 12 (the one that brings translational GTPases to the ribosome), has a double life. Apart from doing its normal job as a part of the ribosome, it doubles as a transctiptional factor, selectively associating with human mitochondrial RNA polymerase and activating it (Surovtseva et al. PNAS 2011). And as if it is not enough, there are several paralogues of L7 12 in mitochondria, both in plants (Delage et al. Biochimie 2007) and in mammals (Koc et al. JBC 2001).
But this seems not tight enough interaction for mitochondrial translation and transcription. It turnes out what mitohondrial ribosomal protein L7 12 (the one that brings translational GTPases to the ribosome), has a double life. Apart from doing its normal job as a part of the ribosome, it doubles as a transctiptional factor, selectively associating with human mitochondrial RNA polymerase and activating it (Surovtseva et al. PNAS 2011). And as if it is not enough, there are several paralogues of L7 12 in mitochondria, both in plants (Delage et al. Biochimie 2007) and in mammals (Koc et al. JBC 2001).
Monday, March 12, 2012
Measuring nucleotide concentrations inside the living cells
Taking biological system apart and doing experiments in vitro is a very powerful approach. However, Nature has loads of dirty tricks up her sleeve, so doing experiments in vivo is more kosher - at least you get all the concentrations rights and will have all of the components present in the system.
Paige JS, Nguyen-Duc T, Song W, & Jaffrey SR (2012). Fluorescence imaging of cellular metabolites with RNA. Science (New York, N.Y.), 335 (6073) PMID: 22403384
Cells use a whole plethora of nucleotide-based messengers (Pesavento and Hengge, Curr. Opin. Microbiol. 2009), and following concentrations of these in vivo is something microbiologists would love to do. It is possible for some, and c-di-GMP is an example. This nucleotide binds to numerous targets, and one of them is PilZ proteins. When binding to PilZ domain, c-di-GMP promotes a massive structural rearrangement, and this interaction can be monitored by adding a FRET pair to PilZ (Benach et al. EMBO J 2007) (Fig. 1). FRET response can be converted in c-di-GMP concentration using a calibration curve, and - viola! - c-di-GMP can be measured in the individual live cells in real time using a PilZ-GFP-based FRET detector (Christen et al. Science 2010).

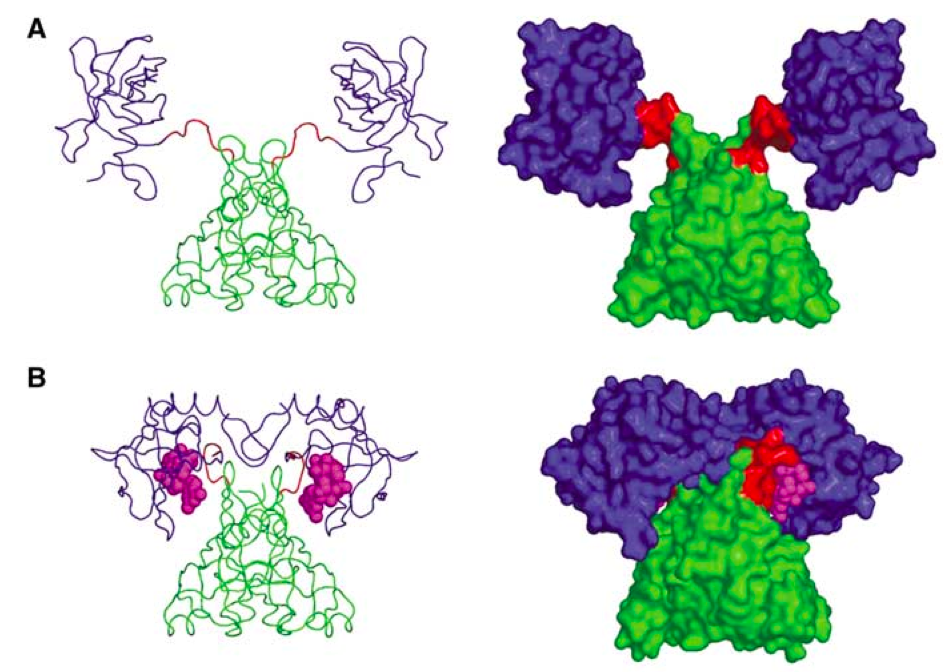
Fig. 1. PlzD: Apo (A) and in complex with c-di-GMP (figure from Benach et al. EMBO J 2007).
The problem with this approach is that is far from being universal. First, one has to have a protein that binds your target nucleotide and undergoes massive rearrangements. Second, this protein should be nice enough to work with so that you can add two GFP molecules to it to make a FRET pair, and still be able to express the protein for in vitro work (one needs to calibrate the FRET response, right?). In the case of stringent response there seem to be no such proteins for detection of my favorite nucleotide, ppGpp... too bad!
The problem with this approach is that is far from being universal. First, one has to have a protein that binds your target nucleotide and undergoes massive rearrangements. Second, this protein should be nice enough to work with so that you can add two GFP molecules to it to make a FRET pair, and still be able to express the protein for in vitro work (one needs to calibrate the FRET response, right?). In the case of stringent response there seem to be no such proteins for detection of my favorite nucleotide, ppGpp... too bad!
Well, there seems to be a new method out there, and this one holds great promise. In their recent Science paper Paige and colleagues use an RNA-based FRET pair using RNA mimic of GFP (Paige et al. Science 2011) combined with a small-molecule-specific aptamer (Fig. 2). When the ligand binds, RNA forms a stable structure and FRET is on! They have followed in E. coli concentrations of two molecules - ADP and SAM. However, aptamers can be evolved for other targets, and this makes this method potentially applicable for detecting whatever molecule that picks your fancy.

References:

Fig. 2. Schematic representartion of the aptamer-based FRET sensor for in vivo detection of small molecules (figure from Paige et al. Science 2012).
References:
Matthias Christen, Hemantha D Kulasekara, Beat Christen, Bridget R Kulasekara, Lucas R Hoffman, and Samuel I Miller (2010) Asymmetrical distribution of the second messenger c-di-GMP upon bacterial cell division. Science (New York, N.Y.), 328 (5983), 1295-7 PMID: 20522779
Benach J, Swaminathan SS, Tamayo R, Handelman SK, Folta-Stogniew E, Ramos JE, Forouhar F, Neely H, Seetharaman J, Camilli A, & Hunt JF (2007). The structural basis of cyclic diguanylate signal transduction by PilZ domains. The EMBO journal, 26 (24), 5153-66 PMID: 18034161
Wednesday, January 4, 2012
PhD student in Molecular Evolution with Dr. Gemma Atkinson
The candidate should have:
- a Masters degree in a biological or computational discipline
- a strong interest in, and enthusiasm for molecular evolution
- familiarity with basic sequence and phylogenetic analyses
- experience in using a programming language such as Python, Perl, Java etc
- fluency in spoken and written English
The PhD will be funded by a monthly stipend, with additional monies available for regular attendance at international conferences and workshops, and for visiting labs abroad. Information on funding is available by request.
Applications should contain:
- a full CV with detailed description of previous relevant experience
- a statement of academic interests
- an electronic version of the Masters thesis
- the names and contact details of at least 2 referees
The candidate is expected to start at the latest September 2012. Please send applications and informal enquiries to gemma.atkinson@ut.ee
More information about the research of Dr Atkinson can be found here.
Update: the call is closed, two applicants selected.
- a Masters degree in a biological or computational discipline
- a strong interest in, and enthusiasm for molecular evolution
- familiarity with basic sequence and phylogenetic analyses
- experience in using a programming language such as Python, Perl, Java etc
- fluency in spoken and written English
The PhD will be funded by a monthly stipend, with additional monies available for regular attendance at international conferences and workshops, and for visiting labs abroad. Information on funding is available by request.
Applications should contain:
- a full CV with detailed description of previous relevant experience
- a statement of academic interests
- an electronic version of the Masters thesis
- the names and contact details of at least 2 referees
The candidate is expected to start at the latest September 2012. Please send applications and informal enquiries to gemma.atkinson@ut.ee
More information about the research of Dr Atkinson can be found here.
Update: the call is closed, two applicants selected.
Subscribe to:
Comments (Atom)