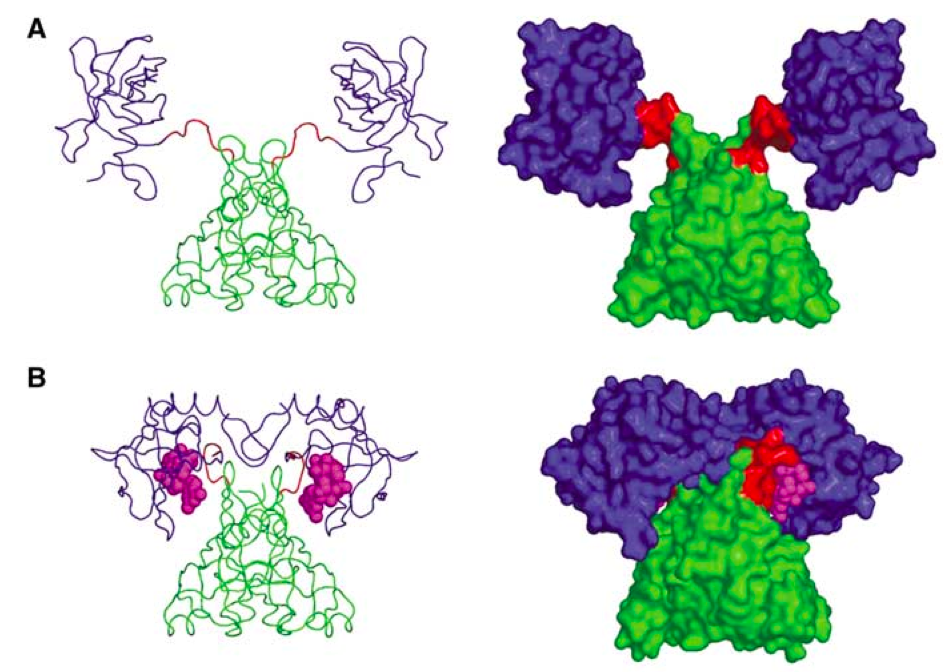
The ribosome is run by translational GTPases. Translational GTPases, in their turn, are regulated by the ribosome. They all bind in the same region (GAC, GTPase associated center) of the ribosome. In bacteria the GTPase binding site consists of a couple of rRNA elements: SRL (sarcin-ricin loop) and thiostrepton loop and several ribosomal proteins:L7/L12 stalk (L10 and L7/L12) and L11.
The latter is the main hero of a fresh paper in Nature Structural and Molecular Biology by Wang and coworkers. They show that bacterial translational GTPases (such as EF-G) when binding to the ribosome act as peptidyl-prolyl cis-trans isomerases (PPIases) driving isomerisation in the conserved residue in the ribosomal protein L11. This isomerisation, in turn, transmits signal to the ribosomal protein L7 /L12 - something that is necessary for efficient GTPase cycling on the ribosome.
I like L11 - it is a key protein for stringent response, and without it stringent factor RelA does not work, as was shown using E. coli mutants lacking L11 (Dabbs J. Bac 1979). These mutants are perfectly viable, but grow ten times slower then the wild type E. coli, most probably due to defects in the ribosome assembly (Hampl et al. JBC 1981). The very viability of the L11 knock-out strains tells us that L11 is not the key for keeping the ribosome running. In fact, less than a half of ribosomal proteins can be knocked-out in E. coli (22 out of 54, Shoji et al. JMB 2011), making L11 one of the less-important ones... and keeping an eye of the translational GTPases is definitely not one of the less-important functions!
This seems to be bit paradoxical - a ribosomal protein that is dispensable involved in something that is very central for protein biosynthesis. It gets even more fascinating when you look at the evolutionary aspect of the story (Gem Atkinson does that in her blog). Wang and colleagues managed to map the PPIase site of EF-G. As they show PPIase activity is universal for all the bacterial translational GTPases they tested, and the PPIase site is, surprisingly, quite a variable region of the G domain! So, do they all reinvent the weel separately? This is all most peculiar.
References:
Wang et al. A conserved proline switch on the ribosome facilitates the recruitment and binding of trGTPases. Nat Struct Mol Biol (2012) PIMD: 22407015The latter is the main hero of a fresh paper in Nature Structural and Molecular Biology by Wang and coworkers. They show that bacterial translational GTPases (such as EF-G) when binding to the ribosome act as peptidyl-prolyl cis-trans isomerases (PPIases) driving isomerisation in the conserved residue in the ribosomal protein L11. This isomerisation, in turn, transmits signal to the ribosomal protein L7 /L12 - something that is necessary for efficient GTPase cycling on the ribosome.
I like L11 - it is a key protein for stringent response, and without it stringent factor RelA does not work, as was shown using E. coli mutants lacking L11 (Dabbs J. Bac 1979). These mutants are perfectly viable, but grow ten times slower then the wild type E. coli, most probably due to defects in the ribosome assembly (Hampl et al. JBC 1981). The very viability of the L11 knock-out strains tells us that L11 is not the key for keeping the ribosome running. In fact, less than a half of ribosomal proteins can be knocked-out in E. coli (22 out of 54, Shoji et al. JMB 2011), making L11 one of the less-important ones... and keeping an eye of the translational GTPases is definitely not one of the less-important functions!
This seems to be bit paradoxical - a ribosomal protein that is dispensable involved in something that is very central for protein biosynthesis. It gets even more fascinating when you look at the evolutionary aspect of the story (Gem Atkinson does that in her blog). Wang and colleagues managed to map the PPIase site of EF-G. As they show PPIase activity is universal for all the bacterial translational GTPases they tested, and the PPIase site is, surprisingly, quite a variable region of the G domain! So, do they all reinvent the weel separately? This is all most peculiar.
References: