Taking biological system apart and doing experiments in vitro is a very powerful approach. However, Nature has loads of dirty tricks up her sleeve, so doing experiments in vivo is more kosher - at least you get all the concentrations rights and will have all of the components present in the system.
Paige JS, Nguyen-Duc T, Song W, & Jaffrey SR (2012). Fluorescence imaging of cellular metabolites with RNA. Science (New York, N.Y.), 335 (6073) PMID: 22403384
Cells use a whole plethora of nucleotide-based messengers (Pesavento and Hengge, Curr. Opin. Microbiol. 2009), and following concentrations of these in vivo is something microbiologists would love to do. It is possible for some, and c-di-GMP is an example. This nucleotide binds to numerous targets, and one of them is PilZ proteins. When binding to PilZ domain, c-di-GMP promotes a massive structural rearrangement, and this interaction can be monitored by adding a FRET pair to PilZ (Benach et al. EMBO J 2007) (Fig. 1). FRET response can be converted in c-di-GMP concentration using a calibration curve, and - viola! - c-di-GMP can be measured in the individual live cells in real time using a PilZ-GFP-based FRET detector (Christen et al. Science 2010).

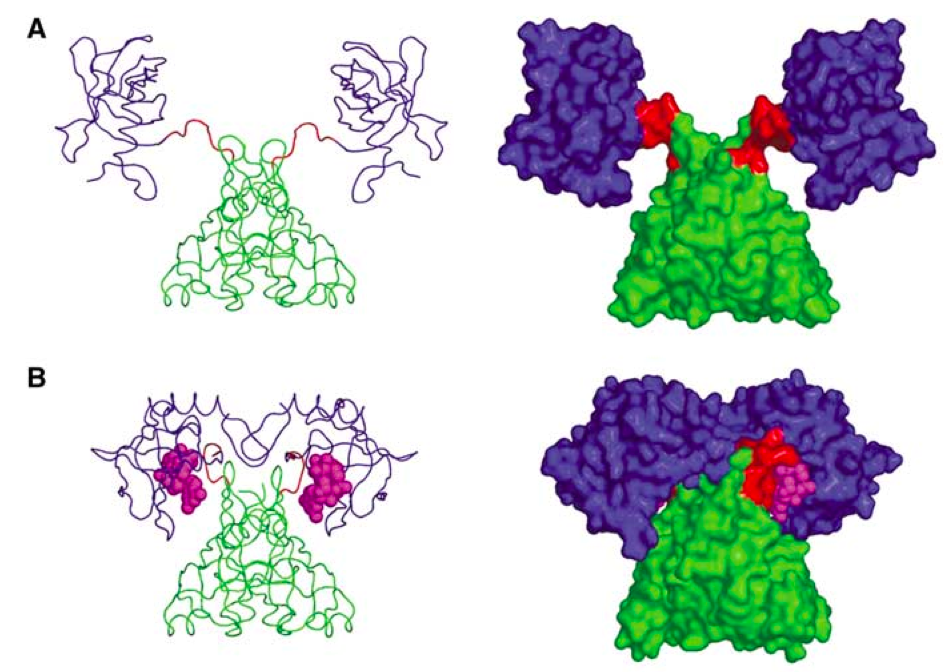
Fig. 1. PlzD: Apo (A) and in complex with c-di-GMP (figure from Benach et al. EMBO J 2007).
The problem with this approach is that is far from being universal. First, one has to have a protein that binds your target nucleotide and undergoes massive rearrangements. Second, this protein should be nice enough to work with so that you can add two GFP molecules to it to make a FRET pair, and still be able to express the protein for in vitro work (one needs to calibrate the FRET response, right?). In the case of stringent response there seem to be no such proteins for detection of my favorite nucleotide, ppGpp... too bad!
The problem with this approach is that is far from being universal. First, one has to have a protein that binds your target nucleotide and undergoes massive rearrangements. Second, this protein should be nice enough to work with so that you can add two GFP molecules to it to make a FRET pair, and still be able to express the protein for in vitro work (one needs to calibrate the FRET response, right?). In the case of stringent response there seem to be no such proteins for detection of my favorite nucleotide, ppGpp... too bad!
Well, there seems to be a new method out there, and this one holds great promise. In their recent Science paper Paige and colleagues use an RNA-based FRET pair using RNA mimic of GFP (Paige et al. Science 2011) combined with a small-molecule-specific aptamer (Fig. 2). When the ligand binds, RNA forms a stable structure and FRET is on! They have followed in E. coli concentrations of two molecules - ADP and SAM. However, aptamers can be evolved for other targets, and this makes this method potentially applicable for detecting whatever molecule that picks your fancy.

References:

Fig. 2. Schematic representartion of the aptamer-based FRET sensor for in vivo detection of small molecules (figure from Paige et al. Science 2012).
References:
Matthias Christen, Hemantha D Kulasekara, Beat Christen, Bridget R Kulasekara, Lucas R Hoffman, and Samuel I Miller (2010) Asymmetrical distribution of the second messenger c-di-GMP upon bacterial cell division. Science (New York, N.Y.), 328 (5983), 1295-7 PMID: 20522779
Benach J, Swaminathan SS, Tamayo R, Handelman SK, Folta-Stogniew E, Ramos JE, Forouhar F, Neely H, Seetharaman J, Camilli A, & Hunt JF (2007). The structural basis of cyclic diguanylate signal transduction by PilZ domains. The EMBO journal, 26 (24), 5153-66 PMID: 18034161
No comments:
Post a Comment